
Abstract
Monocotyledons are distinguishable from dicotyledons by their subtype P2 sieve-element plastids containing cuneate protein crystals, a synapomorphic character uniformly present from basal groups through Lilioids to Commelinoids. The dicotyledon generaAsarum andSaruma (Aristolochiaceae-Asaroideae) are the only other taxa with cuneate crystals, but their sieveelement plastids include an additional large polygonal crystal, as is typical of many eumagnoliids. New investigations in Melanthiaceae s.l. revealed the same pattern (polygonal plus cuneate crystals) in the sieve-element plastids ofJaponolirion osense (Japonoliriaceae/Petrosaviaceae), ofHarperocallis flava, Pleea tenuifolia, andTofleldia (all: Tofieldiaceae). InNarthecium ossifragum a large crystal, present in addition to cuneate ones, usually breaks up into several small crystals, whereas inAletris glabra andLophiola americana (Nartheciaceae) and in all of the 15 species studied and belonging to Melanthiaceae s.str. only cuneate crystals are found. Highresolution TEM pictures reveal a crystal substructure that is densely packed in both cuneate and polygonal forms, but in Tofieldiaceae the polygonal crystals stain less densely, probably as a result of the slightly wider spacing of their subunits. The small crystals ofNarthecium are “loose”; that is, much more widely spaced. Such “loose” crystals are commonly found in sieve-element plastids of Velloziaceae, present there in addition to angular crystals, and together with cuneate crystals in a few Lilioids and many taxa of Poales (Commelinoids). Ontogenetic studies of the sieve elements ofSaruma, Aristolochia, and several monocotyledons have shown that in their plastids cuneate crystals develop very early and independent from a polygonal one present in some taxa. Therefore, a conceivable particulation of polygonal into cuneate crystals is excluded. Consequently, mutations of some monocotyledons that contain a lone, large, polygonal crystal in their sieve-element plastids are explained as the result of a complex genetic block. The total result of all studies in sieve-element plastids suggests thatJaponolirion and Tofieldiaceae are the most basal monocotyledons and that Aristolochiaceae are their dicotyledon sister group.
Zusammenfassung
Die Siebelementplastiden der Monokotyledonen unterscheiden sich von denen aller anderen Samenpflanzen durch keilförmige Proteinkristalle, ein synapomorphes Merkmal, das in Vertretern aller höheren Taxa vorhanden ist.Asarum undSaruma (Aristolochiaceae-Asaroideae) sind die einzigen Vertreter außerhalb der Monokotylen, die ebenfalls dieses Merkmal besitzen. Ihre Siebelementplastiden enthalten aber daneben auch einen großen polygonalen Kristall, wie er bei vielen Eumagnoliiden angetroffen wird. In Untersuchungen an Vertretern der Melanthiaceae s.l. wurde das gleiche Muster in Piastiden vonJaponolirion osense (Japonoliriaceae/ Petrosaviaceae),Harperocallis flava, Pleea tenuifolia und mehrerenTofieldia-Arten (alle: Tofieldiaceae) gefunden. Die Siebelementplastiden vonNarthecium ossifragum enthalten neben keilförmigen Kristallen ebenfalls einen großen Kristall, der aber in der Regel in mehrere kleinere zerfällt, während inAletris glabra undLophiola americana (Nartheciaceae) und 15 weiteren Arten, die jetzt zu den Melanthiaceae s.str. gezählt werden, nur keilförmige Kristalle gefunden wurden. In elektronenmikroskopischen Abbildungen mit hoher Auflösung erscheinen die keilförmigen und polygonalen Kristalle als aus dicht gepackten Untereinheiten zusammengesetzt. Allerdings sind die polygonalen Kristalle der Tofieldiaceae meist weniger stark kontrastiert, vermutlich als Ergebnis einer etwas lockeren Anordnung ihrer Untereinheiten. Die kleinen Kristalle vonNarthecium haben eine weitaus lockere Zusammensetzung, d.h. ihre Untereinheiten stehen in weiterem Abstand. Solche ‚’lockeren” Kristalle sind auch in den Plastiden anderer Taxa anzutreffen: in den Velloziaceae regelmäßig neben angularen Kristallen und in einigen Vertretern der Lilianae und Commelinanae (z.B. in vielen Poales) neben keilförmigen Kristallen. Untersuchungen zur Ontogenie der Siebelemente vonSaruma, Aristolochia und mehreren Monokotyledonen verdeutlichen, dass keilförmige Kristalle sehr früh in der Differenzierung der Piastiden gebildet werden und auch unabhängig von einem eventuell vorher schon vorhandenen polygonalem Kristall. Es wird daher ausgeschlossen, dass keilförmige Kristalle durch einen partiellen oder vollständigen Zerfall von polygonalen entstehen. Dementsprechend sind auch die großen polygonalen Kristalle, die als alleiniger Einschluss von Siebelementplastiden in Mutationen weniger Monokotylen gefunden wurden, nicht als Vorläufer von keilförmigen Kristallen, sondern als durch einen komplexen genetischen Block entstanden, zu erklären. In Summe der Untersuchungen an Siebelementplastiden erscheinenJaponolirion und die Tofieldiaceae als an der Basis der Monokotyledonen stehende Taxa und die Aristolochiaceae als ihre Schwestergruppe unter den Dicotyledonen.
Similar content being viewed by others


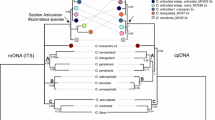
Literature Cited
Ambrose, J. D. 1980. A re-evaluation of the Melanthioideae (Liliaceae) using numerical analyses. Pp. 65–81in C. D. Brickell, D. F. Cutler & M. Gregory (eds.), Petaloid monocotyledons: Horticultural and botanical research. Linn. Soc. Symposium Ser., 8. Academic Press, London, New York.
APG (Angiosperm Phylogeny Group). 1998. An ordinal classification for the families of flowering plants. Ann. Missouri Bot. Gard. 85: 531–553.
Behnke, H.-D. 1971. Zum Feinbau der Siebröhren-Plastiden vonAristolochia undAsarum (Aristolochiaceae). Planta 97: 62–69.
—. 1972. Sieve-tube plastids in relation to angiosperm systematics: An attempt towards a classification by ultrastructural analysis. Bot. Rev. (Lancaster) 38: 155–197.
—. 1974. Sieve-element plastids of Gymnospermae: Their ultrastructure in relation to systematics. Pl. Syst. Evol. 123: 1–12.
—. 1981a. Siebelement-Plastiden, Phloem-Protein und Evolution der Blütenpflanzen, II. Monokotyledonen. Ber. Deutsch. Bot. Ges. 94: 647–662.
—. 1981b. Sieve-element characters. Nord. J. Bot. 1: 381–400.
—. 1988. Sieve-element plastids, phloem protein and evolution of flowering plants, III. Magnoliidae. Taxon 37, 699–732.
—. 1990. Sieve elements of internodal and nodal anastomoses of the monocotyledon lianaDioscorea. Pp. 161–178in H.-D. Behnke & R. D. Sjolund (eds.), Sieve elements: Comparative structure, induction and development. Springer-Verlag, Berlin, Heidelberg, New York.
—. 1991. Distribution and evolution of forms and types of sieve-element plastids in the dicotyledons. Aliso 13: 167–182.
—. 1995. P-type sieve-element plastids and the systematics of the Arales (sensu Cronquist 1988), with S-type plastids inPistia. Pl. Syst. Evol. 195: 87–119.
—. 2000. Forms and sizes of sieve-element plastids and evolution of the monocotyledons. Pp. 163–188in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. CSIRO, Melbourne, Australia.
—. 2000. Systematics and evolution of the Velloziaceae, with special reference to sieve-element plastids andrbcL sequence data. Bot. J. Linn. Soc. 134: 93–129.
Bessey, C. E. 1893. Evolution and classification. Bot. Gaz. 18: 329–333.
Caddick, L. R., P. J. Rudall, P. Wilkin &M. W. Chase. 2000. Yams and their allies: Systematics of Dioscoreales. Pp. 475–487in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. CSIRO, Melbourne, Australia.
Chase, M. W., D. E. Soltis, R. G. Olmstead, D. Morgan, D. H. Les, B. D. Mishler, M. R. Duvall, R. A. Price, H. G. Hills, Y.-L. Qiu, K. A. Kron, J. H. Rettig, E. Conti, J. D. Palmer, J. R. Manhart, K. J. Sytsma, H. J. Michaels, W. J. Kress, K. G. Karol, W. D. Clark, M. Hedén, B. S. Gaut, R. K. Jansen, K.-J. Kim, C. F. Wimpee, J. F. Smith, G. R. Furnier, S. H. Strauss, Q.-Y. Xiang, G. M. Plunkett, P. S. Soltis, S. M. Swensen, S. E. Williams, P. A. Gadek, C. J. Quinn, L. E. Eguiarte, E. Golenberg, G. H. Learn Jr., S. W. Graham, S. C. H. Barrett, S. Dayanandan &V. A. Albert. 1993. Phylogenetics of seed plants: An analysis of nucleotide sequences from the plastid generbcL. Ann. Missouri Bot. Gard. 80: 528–580.
—. 1995a. Molecular phylogenetics of Lilianae. Pp. 109–137in P. J. Rudall, P. J. Cribb, D. F. Cutler & C. J. Humphries (eds.), Monocotyledons: Systematics and evolution. 2 vols. Roy. Bot. Gard., Kew.
—. 1995b. Monocot systematics: A combined analysis. Pp. 685–730in P. J. Rudall, P. J. Cribb, D. F. Cutler & C. J. Humphries (eds.), Monocotyledons: Systematics and evolution. 2 vols. Roy. Bot. Gard., Kew.
—. 2000. Higher-level systematics of the monocotyledons: An assessment of current knowledge and a new classification. Pp. 3–16in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. CSIRO, Melbourne, Australia.
Cronquist, A. 1981. An Integrated System of Classification of Flowering Plants. Columbia Univ. Press, New York.
—. 1988. The Evolution and Classification of Flowering Plants. Ed. 2. New York Bot. Gard., Bronx.
Dahlgren, R. &K. Bremer. 1985. Major clades of the angiosperms. Cladistics 1: 349–368.
—. 1983. Monocotyledon evolution. Characters and phylogenetic estimation. Evol. Biol. 16: 255–395.
—. 1985. The families of the monocotyledons: Structure, evolution, and taxonomy. Springer-Verlag, Berlin.
Duvall, M. R. 2000. Seeking the sister group of the monocots. Pp. 25–32in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. CSIRO, Melbourne, Australia.
—. 1993a. Phylogenetic analysis ofrbcL sequences identifiesAcorus calamus as the primal extant monocotyledon. Proc. Natl. Acad. U.S.A. 90: 4641–4644.
—. 1993b. Phylogenetic hypotheses for the monocotyledons constructed fromrbcL sequence data. Ann. Missouri Bot. Gard. 80: 607–619.
Engler, A. 1892. Die systematische Anordnung der monokotyledonen Angiospermen. Abh. Preuss. Akad. Wiss.
Fuse, S. &M. N. Tamura. 2000. A phylogenetic analysis of the plastid matK gene with emphasis on Melanthiaceae sensu lato. Pl. Biol. 2: 415–427.
Goldblatt, P. 1995. The status of R. Dahlgren’s orders Liliales and Melanthiales. Pp. 181–200in P. J. Rudall, P. J. Cribb, D. F. Cutler & C. J. Humphries (eds.), Monocotyledons: Systematics and evolution. 2 vols. Roy. Bot. Gard., Kew.
Grayum, M. H. 1987. A summary of evidence and arguments supporting the removal ofAcorus from the Araceae. Taxon 36: 723–729.
Hallier, H. 1905. Ein zweiter Entwurf des natürlichen (phylogenetischen) Systems der Blutenpflanzen. Vorläufige Mitteilung. Ber. Deutsch. Bot. Ges. 23: 85–91.
Holmgren, P. K., N. H. Holmgren &L. C. Barnett (eds.). 1990. Index herbariorum, I. The herbaria of the world. Ed. 8. New York Bot. Gard., Bronx.
Huber, H. 1969. Die Samenmerkmale und Verwandtschaftsverhältnisse der Liliifloren. Mitt. Bot. Staatssamml. München 8: 219–538.
—. 1977. The treatment of the monocotyledons in an evolutionary system of classification. Pl. Syst. Evol., Suppl. 1: 285–296.
—. 1982. Die zweikeimblättrigen Gehölze im System der Angiospermen. Mitt. Bot. Staatssamml. München 18: 59–78.
—. 1991. Angiospermen: Leitfaden durch die Ordnungen und Familien der Bedecktsamer. Fischer-Verlag, Stuttgart, New York.
Kubitzki, K., P. J. Rudall &M. W. Chase. 1998. Systematics and Evolution. Pp. 23–33in K. Kubitzki (ed.), The families and genera of vascular plants. Vol. 3. Flowering plants: Monocotyledons: Lilianae (except Orchidaceae). Springer-Verlag, Berlin.
Nakai, T. 1930. Plantae Japonicae and Koreanae. Bot. Mag. (Tokyo) 44: 22–23.
Qiu, Y.L., J. Lee, F. Bernasconi-Quadroni, D. E. Soltis, P. S. Soltis, M. Zanis, E. A. Zimmer, Z. Chen, V. Savolainen &M. W. Chase. 2000. Phylogeny of basal angiosperms: analyses of five genes from three genomes. Int. J. Pl. Sci. 161 (6 Suppl.): S3-S27.
Rudall, P. J., P. J. Cribb, D. F. Cutler &C. J. Humphries (eds.). 1995. Monocotyledons: Systematics and evolution. 2 vols. Roy. Bot. Gard., Kew.
—. 2000. Consider the lilies: Systematics of the Liliales. Pp. 347–359in K. L. Wilson & D. A. Morrison (eds.), Monocots: Systematics and evolution. CSIRO, Melbourne, Australia.
Savolainen, V., M. F. Fay, D. C. Albach, A. Backlund, M. van der Bank, K. M. Cameron, S. A. Johnson, M. D. Lledo, J.-C. Pintaud, M. Powell, M. C. Sheahan, D. E. Soltis, P. S. Soltis, P. Weston, W. M. Whitten, K. J. Wurdack &M. W. Chase. 2000. Phylogeny of the eudicots: A nearly complete familial analysis based onrbcL gene sequences. Kew Bull. 55: 257–309.
Soltis, D. E., P. S. Soltis, M. W. Chase, M. E. Mort, D. C. Albach, M. Zanis, V. Savolainen, W. H. Hahn, S. B. Hoot, M. F. Fay, M. Axtell, S. M. Swensen, L. M. Prince, W. J. Kress, K. C. Nixon &J. S. Farris. 2000. Angiosperm phylogeny inferred from 18S rDNA,rbcL, andatpB sequences. Bot. J. Linn. Soc. 133: 381–461.
Takhtajan, A. 1997. Diversity and classification of flowering plants. Columbia Univ. Press, New York.
Tamura, M. N. 1998. Nartheciaceae. Pp. 381–392in K. Kubitzki (ed.), The families and genera of vascular plants. Vol. 3. Flowering plants: Monocotyledons: Lilianae (except Orchidaceae). Springer-Verlag, Berlin.
Tillich, H.-J. 1985. Keimlingsbau und verwandtschaftliche Beziehungen der Araceae. Gleditschia 13: 63–73.
Utech, F. H. 1984. Floral vascular anatomy ofJaponolirion osense Nakai (Liliaceae) and its tribal relationship. Ann. Carnegie Mus. 53: 447–461.
Walker, J. W. 1986. Classification and evolution of the monocotyledons. Amer. J. Bot. 73: 746.
Wilson, K. L. &D. A. Morrison (eds.). 2000. Monocots: Systematics and evolution. CSIRO, Melbourne, Australia.
Zomlefer, W. B. 1999. Advances in angiosperm systematics: Examples from the Liliales and Asparagales. J. Torrey Bot. Soc. 125: 58–62.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Behnke, H.D. Sieve-element plastids and evolution of monocotyledons, with emphasis on Melanthiaceae sensu lato and Aristolochiaceae-Asaroideae, a putative dicotyledon sister group. Bot. Rev 68, 524–544 (2002). https://doi.org/10.1663/0006-8101(2002)068[0524:SPAEOM]2.0.CO;2
Issue Date:
DOI: https://doi.org/10.1663/0006-8101(2002)068[0524:SPAEOM]2.0.CO;2